Важнейшим компонентом клеточного цикла является митотический (пролиферативный) цикл. Он представляет собой комплекс взаимосвязанных и согласованных явлений во время деления клетки, а также до и после него. Митотический цикл - это совокупность процессов, происходящих в клетке от одного деления до следующего и заканчивающихся образованием двух клеток следующей генерации. Кроме этого, в понятие жизненного цикла входят также период выполнения клеткой своих функций и периоды покоя. В это время дальнейшая клеточная судьба неопределенна: клетка может начать делиться (вступает в митоз) либо начать готовиться к выполнению специфических функций.
Основные стадии митоза
1.
Редупликация (самоудвоение) генетической информации материнской клетки и
равномерное распределение ее между дочерними клетками. Это сопровождается
изменениями структуры и морфологии хромосом, в которых сосредоточено более 90%
информации эукариотической клетки.
2.
Митотический цикл состоит из четырех последовательных периодов: пресинтетического (или постмитотического) G1, синтетического S,
постсинтетического (или премитотического) G2 и собственно митоза. Они составляют
автокаталитическую интерфазу (подготовительный период).
Фазы клеточного цикла:
1) пресинтетическая (G1). Идет сразу после деления клетки. Синтеза ДНК еще не происходит. Клетка активно растет в размерах, запасает вещества, необходимые для деления: белки (гистоны, структурные белки, ферменты), РНК, молекулы АТФ. Происходит деление митохондрий и хлоропластов (т. е. структур, способных к ауторепродукции). Восстанавливаются черты организации интерфазной клетки после предшествующего деления;
2)
синтетическая (S). Происходит удвоение генетического материала путем
репликации ДНК. Она происходит полуконсервативным способом, когда двойная
спираль молекулы ДНК расходится на две цепи и на каждой из них синтезируется комплементарная цепочка.
В итоге образуются две идентичные двойные спирали ДНК, каждая из которых состоит
из одной новой и старой цепи ДНК. Количество наследственного материала
удваивается. Кроме этого, продолжается синтез РНК и белков. Также репликации
подвергается небольшая часть митохонд-риальной ДНК (основная же ее часть
реплицируется в G2 период);
3) постсинтетическая (G2). ДНК уже не синтезируется, но происходит исправление недочетов, допущенных при синтезе ее в S период (репарация). Также накапливаются энергия и питательные вещества, продолжается синтез РНК и белков (преимущественно ядерных).
S и G2 непосредственно связаны с митозом, поэтому их иногда выделяют в отдельный
период - препрофазу.
После этого наступает собственно митоз, который состоит из четырех фаз. Процесс
деления включает в себя несколько последовательных фаз и представляет собой
цикл. Его продолжительность различна и составляет у большинства клеток от 10 до
50 ч. При этом у клеток тела человека продолжительность самого митоза составляет
1-1,5 ч, G2-периода интерфазы - 2-3 ч, S-периода интерфазы - 6-10 ч.
Длительность отдельных стадий различна и варьируется в зависимости от типа
ткани, физиологического состояния организма, внешних факторов. Наиболее
продолжительны стадии сопряженные с процессами внутриклеточного синтеза: профаза
и телофаза. Наиболее быстротечны фазы митоза, в ходе которых происходит движение
хромосом: метафаза и анафаза. Непосредственно процесс расхождения хромосом к
полюсам обычно не превышает 10 минут.
Профаза
К основным событиям профазы относят конденсацию хромосом внутри ядра и
образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе
является характерной, но не обязательной для всех клеток особенностью.
Условно за начало профазы принимается момент возникновения микроскопически
видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение
хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения
сопровождаются повышением активности фосфорилаз, модифицирующих гистоны,
непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается
транскрипционная активность хроматина, инактивируются ядрышковые гены, большая
часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в
ранней профазе остаются спаренными по всей своей длине с помощью
белков-когезинов, однако к началу прометафазы связь между хроматидами
сохраняется лишь в области центромер. К поздней профазе на каждой центромере
сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для
присоединения к микротрубочкам веретена деления в прометафазе.
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено - одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца, микротрубочки и кинетохоры хромосом.
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут до 15 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления.
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.
Прометафаза
Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом. В результате такого взаимодействия кинетохорные микротрубочки стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.
В клетках млекопитающих прометафаза протекает, как правило, в течение 10-20 минут. В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона - около 30 минут.
Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи - к периферии клетки. В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.
В метафазе, также как и в течение других фаз митоза, продолжается активное
обновление микротрубочек веретена путём интенсивной сборки и деполимеризации
молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных
микротрубочек, происходит постоянная переборка межполюсных микротрубочек,
численность которых в метафазе достигает максимума.
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид,
соединение между которыми сохраняется лишь в центромерных участках. Плечи
хроматид располагаются параллельно друг другу, и становится отчетливо заметной
разделяющая их щель.
Анафаза
Анафаза - самая короткая стадия митоза, которая начинается внезапным
разделением и последующим расхождением сестринских хроматид в направлении
противоположных полюсов клетки. Хроматиды расходятся с равномерной скоростью
достигающей 0,5-2 мкм/мин., при этом они часто принимают V-образную форму. Их
движение обусловлено воздействием значительных сил, оценочно 10 дин на
хромосому, что в 10 000 раз превышает усилие, необходимое для простого
продвижения хромосомы через цитоплазму с наблюдаемой скоростью.
Как правило, расхождение хромосом в анафазе состоит из двух относительно
независимых процессов называемых анафазой А и анафазой В.
Анафаза А характеризуется расхождением сестринских хроматид к противоположным
полюсам деления клетки. За их движение при этом отвечают те же силы, что ранее
удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения
хроматид сопровождается сокращением длины деполимеризующихся кинетохорных
микротрубочек. Причем их распад наблюдается преимущественно в области
кинетохоров, со стороны плюс-концов. Вероятно, деполимеризация микротрубочек у
кинетохоров либо в области полюсов деления является необходимым условием для
перемещения сестринских хроматид, так как их движение прекращается при
добавлении таксола или тяжёлой воды, оказывающих стабилизирующее воздействие на
микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока
остается неизвестным.
Во время анафазы В расходятся сами полюса деления клетки, и, в отличии от
анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со
стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при
взаимодействии отчасти и создают расталкивающее полюса усилие. Величина
относительного перемещения полюсов при этом, также как и степень перекрывания
полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей
разных видов. Помимо расталкивающих сил, на полюса деления воздействуют тянущие
силы со стороны астральных микротрубочек, которые создаются в результате
взаимодействия с динеино-подобными белками на плазматической мембране клетки.
Последовательность, продолжительность и относительный вклад каждого из двух
процессов, слагающих анафазу, могут быть крайне различны. Так в клетках
млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к
противоположным полюсам и продолжается вплоть до удлинения митотического
веретена в 1,5-2 раза по сравнению с метафазным. В некоторых других клетках
анафаза В начинается только после того как хроматиды достигают полюсов деления.
У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по
сравнению с метафазным. В растительных клетках анафаза В отсутствует.
Телофаза
Телофаза рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга.
Окончание телофазы преимущественно совпадает с разделением тела материнской
клетки - цитокинезом. При этом образуются две или более дочерние клетки.
Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине
анафазы и могут продолжаться после завершения телофазы. Митоз не всегда
сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в
качестве отдельной фазы митотического деления и обычно рассматривается в составе
телофазы.
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки и
деление путём образования клеточной пластинки. Плоскость деления клетки
детерминируется положением митотического веретена и проходит под прямым углом к
длинной оси веретена.
При делении поперечной перетяжкой клетки место разделения цитоплазмы
закладывается предварительно ещё в период анафазы, когда в плоскости метафазной
пластинки под мембраной клетки возникает сократимое кольцо из актиновых и
миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца,
образуется борозда деления, которая постепенно углубляется вплоть до полного
разделения клетки. По окончании цитокинеза сократимое кольцо полностью
распадается, а плазматическая мембрана стягивается вокруг остаточного тельца
Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек,
тесно упакованных вместе с материалом плотного матрикса.
Деление путём образования клеточной пластинки начинается с перемещения мелких
ограниченных мембраной пузырьков по направлению к экваториальной плоскости
клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру
- раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата
Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных
микротрубочек веретена деления, образующих цилиндрическую структуру, называемую
фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего
фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых
мембранных пузырьков продолжается рост клеточной пластинки вплоть до её
окончательного слияния с мембраной материнской клетки. После окончательного
разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы
целлюлозы, завершая образование жёсткой клеточной стенки.
Для определения завершения каждой фазы клеточного цикла необходимо наличие в нем контрольных точек. Если клетка «проходит» контрольную точку то она продолжается «двигаться» по клеточному циклу. Если же какие-либо обстоятельства, например повреждение ДНК, мешают клетке пройти через контрольную точку, которую можно сравнить со своего рода контрольным пунктом, то клетка останавливается и другой фазы клеточного цикла не наступает по крайней мере до тех пор, пока не будут устранены препятствия, не позволявшие клетке пройти через контрольный пункт.
Рост и развитие живых организмов невозможен без процессов деления клеток. Одним из них является митоз - процесс деления эукариотических клеток, при котором передаётся и сохраняется генетическая информация. В этой статье Вы подробнее узнаете об особенностях митотического цикла, познакомитесь с характеристикой всех фаз митоза, которая будет внесена в таблицу.
Понятие «митотический цикл»
Все процессы, которые происходят в клетке, начиная от одного деления до другого, и заканчивая получением двух дочерних клеток, называется митотическим циклом. Жизненным циклом клетки также является состояние покоя и период выполнения своих прямых функций.
К основным стадиям митоза относятся:
- Самоудвоение или редупликация генетического кода , который передаётся от материнской клетки к двум дочерним. Процесс влияет на структуру и образование хромосом.
- Клеточный цикл - состоит из четырёх периодов: пресинтетического, синтетического, постсинтетического и, собственно, митоза.
Первые три периода (пресинтетический, синтетический и постсинтетический) относятся к интерфазе митоза.
Некоторые учёные синтетический и постсинтетический период называют препрофазой митоза. Так как все стадии происходят непрерывно, плавно переходя от одной к другой, чёткого разделения между ними нет.
Процесс непосредственного деления клетки, митоз, происходит в четыре фазы, соответствуя такой последовательности:
ТОП-4 статьи которые читают вместе с этой
- Профаза;
- Метафаза;
- Анафаза;
- Телофаза.

Рис. 1. Фазы митоза
Познакомиться с кратким описанием каждой фазы можно в таблице «Фазы митоза», которая представлена далее.
Таблица «Фазы митоза»
|
№ п/п |
Фаза |
Характеристика |
|
В профазе митоза происходит растворение ядерной оболочки и ядрышка, центриоли расходятся к разным полюсам, начинается формирование микротрубочек, так называемых нитей веретена деления, конденсируются хроматиды в хромосомах. |
||
|
Метафаза |
На этом этапе максимально конденсируются хроматиды в хромосомах и выстраиваются в экваториальной части веретена, образуя метафазную пластинку. Нити центриолей прикрепляются к центромерам хроматид или растягиваются между полюсами. |
|
|
Является самой кратковременной фазой, во время которой происходит разделение хроматид после распада центромер хромосом. Пара расходится к разным полюсам и начинает самостоятельный образ жизни. |
||
|
Телофаза |
Является заключительным этапом митоза, при котором новообразованные хромосомы обретают обычные размеры. Вокруг них образуется новая ядерная оболочка с ядрышком внутри. Нити веретена распадаются и исчезают, начинается процесс деления цитоплазмы и её органоидов (цитотомия). |
Процесс цитотомии в животной клетке происходит при помощи борозды деления, а в растительной клетке - с помощью клеточной пластинки.
Нетипичные формы митоза
В природе иногда встречаются и нетипичные формы митоза:
- Амитоз - способ прямого деления ядра, при котором сохраняется строение ядра, ядрышко не распадается, хромосомы при этом не просматриваются. В результате получаем двухъядерную клетку.

Рис. 2. Амитоз
- Политения - кратно увеличиваются клетки ДНК, но без увеличения содержания хромосом.
- Эндомитоз - в ходе процесса после репликации ДНК нет разделения хромосом на дочерние хроматиды. При этом число хромосом увеличивается в десятки раз, возникают полиплоидные клетки, которые могут привести к мутации.

Рис. 3. Эндомитоз
Что мы узнали?
Процесс непрямого деления клеток-эукариотов проходит в несколько этапов, каждый из которых имеет свои особенности. Митотический цикл состоит из стадий интерфазы и непосредственного клеточного деления, состоящего из четырёх фаз: профазы, метафазы, анафазы и телофазы. Иногда в природе встречаются нетипичные способы деления, к ним относятся амитоз, политения и эндомитоз.
Тест по теме
Оценка доклада
Средняя оценка: 4.4 . Всего получено оценок: 518.
Деление клетки является центральным моментом размножения.
В процессе деления из одной клетки возникают две. Клетка на основе ассимиляции органических и неорганических веществ создает себе подобную с характерным строением и функциями.
В делении клетки можно наблюдать два основных момента: деление ядра - митоз и деление цитоплазмы - цитокинез, или цитотомия. Основное внимание генетиков до сих пор приковывает митоз, поскольку, с точки зрения хромосомной теории, ядро считается «органом» наследственности.
В процессе митоза происходит:
- удвоение вещества хромосом;
- изменение физического состояния и химической организации хромосом;
- расхождение дочерних, точнее сестринских, хромосом к полюсам клетки;
- последующее деление цитоплазмы и полное восстановление двух новых ядер в сестринских клетках.
Таким образом, в митозе заложен весь жизненный цикл ядерных генов: удвоение, распределение и функционирование; в результате завершения митотического цикла сестринские клетки оказываются с равным «наследством».
При делении ядро клетки проходит пять последовательных стадий: интерфазу, профазу, метафазу, анафазу и телофазу; некоторые цитологи выделяют еще шестую стадию - прометафазу.
Между двумя последовательными делениями клетки ядро находится в стадии интерфазы. В этот период ядро при фиксации и Окраске имеет сетчатую структуру, образуемую красящимися тонкими нитями, которые в следующей фазе формируются в хромосомы. Хотя интерфазу называют иначе фазой покоящегося ядра , на самом теле метаболические процессы в ядре в этот период совершаются с наибольшей активностью.
Профаза - первая стадия подготовки ядра к делению. В профазе сетчатая структура ядра постепенно превращается в хромосомные нити. С самой ранней профазы даже в световом микроскопе можно наблюдать двойную природу хромосом. Это говорит о том, что в ядре именно в ранней или поздней интерфазе осуществляется наиболее важный процесс митоза - удвоение, или редупликация, хромосом, при котором каждая из материнских хромосом строит себе подобную - дочернюю. Вследствие этого каждая хромосома выглядит продольно удвоенной. Однако эти половинки хромосом, которые называются сестринскими хроматидами , в профазе не расходятся, так как удерживаются вместе одним общим участком - центромерой; центромерный участок делится позже. В профазе хромосомы претерпевают процесс скручивания по своей оси, что приводит к их укорочению и утолщению. Нужно подчеркнуть, что в профазе каждая хромосома в кариолимфе располагается случайно.
В клетках животных еще в поздней телофазе или очень ранней интерфазе происходит удвоение центриоли, после чего в профазе начинается схождение дочерних центриолей к полюсам и образований астросферы и веретена, называемого новым аппаратом. В это же время растворяются ядрышки. Существенным признаком окончания профазы является растворение оболочки ядра, в результате чего хромосомы оказываются в общей, массе цитоплазмы и кариоплазмы, которые теперь образуют миксоплазму. Этим заканчивается профаза; клетка вступает в метафазу.
В последнее время между профазой и метафазой исследователи стали выделять промежуточную стадию, называемую прометафазой . Прометафаза характеризуется растворением и исчезновением ядерной оболочки и движением хромосом к экваториальной плоскости клетки. Но к этому моменту еще не завершается образование ахроматинового веретена.
Метафазой называют стадию окончания расположения хромосом на экваторе веретена. Характерное расположение хромосом в экваториальной плоскости называют экваториальной, или метафазной, пластинкой. Расположение хромосом по отношению друг к другу является случайным. В метафазе хорошо выявляются число и форма хромосом, в особенности при рассмотрении экваториальной пластинки с полюсов деления клетки. Ахроматиновое веретено полностью сформировано: нити веретена приобретают плотную консистенцию чем остальная масса цитоплазмы, и прикрепляются к центромерному участку хромосомы. Цитоплазма клетки в этот период имеет наименьшую вязкость.
Анафазой называют следующую фазу митоза, в которой делятся хроматиды, которые теперь можно назвать уже сестринскими или дочерними хромосомами, расходятся к полюсам. При этом отталкиваются друг от друга в первую очередь центромерные участки, а затем расходятся к полюсам сами хромосомы. Нужно сказать, что расхождение хромосом в анафазе начинается одновременно - «как по команде» - и завершается очень быстро.
В телофазе дочерние хромосомы деспирализуются и утрачивают видимую индивидуальность. Образуются оболочка ядра и само ядро. Ядро реконструируется в обратном порядке по сравнению с теми изменениями, которые оно претерпевало в профазе. В конце концов восстанавливаются и ядрышки (или ядрышко), причем в том количестве, в каком они присутствовали в родительских ядрах. Число ядрышек является характерным для каждого типа клеток.
В это же время начинается симметричное разделение тела клетки. Ядра же дочерних клеток переходят в состояние интерфазы.

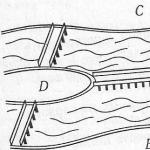
Нa рисунке выше приведена схема цитокинеза животной и растительной клеток. В животной клетке деление происходит путем перешнуровывания цитоплазмы материнской клетки. В растительной клетке формирование клеточной перегородки идет при участки бляшек веретена, образующих в плоскости экватора перегородку, называемую фрагмопластом. Этим заканчивается митотический цикл. Продолжительность его зависит, по-видимому, от типа ткани, физиологического состояния организма, внешних факторов (температуры, светового режима) и длится от 30 мин до 3 ч. По данным разных авторов, скорость прохождения отдельных фаз изменчива.
Как внутренние, так и внешние факторы среды, действующие на рост организма и его функциональное состояние, влияют на продолжительность клеточного деления и его отдельных фаз. Поскольку ядро играет огромную роль в метаболических процессах клетки, естественно полагать, что длительность фаз митоза может изменяться в соответствии с функциональным состоянием ткани органа. Например, установлено, что во время покоя и сна животных митотическая активность различных тканей значительно выше, чем в период бодрствования. У ряда животных частота клеточных делений на свету снижается, а в темноте увеличивается. Предполагают также, что на митотическую активность клетки влияют гормоны.
Причины, определяющие готовность клетки к делению, до сих пор остаются невыясненными. Есть основания предполагать несколько таких причин:
- удвоение массы клеточной протоплазмы, хромосом и других органелл, в силу чего нарушаются ядерно-плазменные отношения; для деления клетка должна достигнуть определенных веса и объема, характерных для клеток данной ткани;
- удвоение хромосом;
- выделение хромосомами и другими органеллами клетки специальных веществ, стимулирующих клеточное деление.
Механизм расхождения хромосом к полюсам в анафазе митоза также остается невыясненным. Активную роль в этом процессе, видимо, играют нити веретена, представляющие организованные и ориентированные центриолями и центромерами белковые нити.
Характер митоза, как мы уже говорили, меняется в зависимости от типа и функционального состояния ткани. Для клеток разных тканей характерны различные типы митозов, В описанном типе митоза деление клетки происходит равным и симметричным образом. В результате симметричного митоза сестринские клетки являются наследственно равноценными в отношении как ядерных генов, так и цитоплазмы. Однако, кроме симметричного, встречаются и другие типы митоза, а именно: асимметричный митоз, митоз с задержкой цитокинеза, деление многоядерных клеток (деление синцитиев), амитоз, эндомитоз, эндорепродукция и политения.
В случае асимметричного митоза сестринские клетки оказываются неравноценными по размеру, количеству цитоплазмы, а также в отношении их дальнейшей судьбы. Примером этого могут служить неодинакового размера сестринские (дочерние) клетки нейробласта кузнечика, яйцеклетки животных при созревании и при спиральном дроблении; при делении ядер в пыльцевых зернах одна из дочерних клеток может в дальнейшем делиться, другая - нет, и т. д.
Митоз с задержкой цитокинеза характеризуется тем, что ядро клетки делится многократно, и лишь затем происходит деление тела клетки. В результате такого деления образуются многоядерные клетки вроде синцития. Примером этого служит образование клеток эндосперма и образование спор.
Амитозом называют прямое деление ядра без образования фигур деления. При этом деление ядра происходит путем «перешнуровывания» его на две части; иногда из одного ядра образуется сразу несколько ядер (фрагментация). Амитоз постоянно встречается в клетках ряда специализированных и патологических тканей, например в раковых опухолях. Его можно наблюдать при воздействиях различных повреждающих агентов (ионизирующие излучения и высокая температура).
Эндомитозом называют такой процесс, когда происходит удвоение деления ядер. При этом хромосомы, как и обычно, репродуцируются в интерфазе, но последующее расхождение их происходит внутри ядра с сохранением ядерной оболочки и без образования ахроматинового веретена. В некоторых случаях хотя и растворяется оболочка ядра, однако расхождение хромосом к полюсам не осуществляется, вследствие чего в клетке происходит умножение числа хромосом даже в несколько десятков раз. Эндомитоз встречается в клетках различных тканей как растений, так и животных. Так, например, А. А. Прокофьева-Бельговская показала, что путем эндомитоза в клетках специализированных тканей: в гиподерме циклопа, жировом теле, перитонеальном эпителии и других тканях кобылки (Stenobothrus) - набор хромосом может увеличиваться в 10 раз. Такое умножение числа хромосом связано с функциональными особенностями дифференцированной ткани.
При политении происходит умножение числа хромосомных нитей: после редупликации по всей длине они не расходятся и остаются прилегающими друг к другу. В этом случае умножается число хромосомных нитей в пределах одной хромосомы, в результате диаметр хромосом заметно увеличивается. Число таких тонких нитей в политенной хромосоме может достигать 1000-2000. В этом случае образуются так называемые гигантские хромосомы. При политении выпадают все фазы митотического цикла, кроме основной - репродукции первичных нитей хромосомы. Явление политении наблюдается в клетках ряда дифференцированных тканей, например в ткани слюнных желез двукрылых, в клетках некоторых растений и простейших.
Иногда имеет место удвоение одной или нескольких хромосом без каких-либо преобразований ядра - такое явление называется эндорепродукцией .
Итак, все фазы митоза клетки, составляющие , являются обязательными лишь для типичного процесса.
некоторых случаях, главным образом в дифференцированных тканях, митотический цикл претерпевает изменения. Клетки таких тканей утратили способность к воспроизведению целого организма, и метаболическая деятельность их ядра приспособлена к функции поциализированной ткани.
Эмбриональные и меристемные клетки, не утратившие функцию воспроизведения целого организма и относящиеся к недифференцированным тканям, сохраняют полный цикл митоза, на чем и основывается бесполое и вегетативное размножение.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Размножение клеток – один из важнейших биологических процессов, является необходимым условием существования всего живого. Репродукция осуществляется путем деления исходной клетки.
Клетка – это наименьшая морфологическая единица строения любого живого организма, способная к самопроизводству и саморегуляции. Время ее существования от деления до гибели или же последующей репродукции называется клеточным циклом.
Ткани и органы состоят из различных клеток, которые имеют свой период существования. Каждая из них растет и развивается, чтобы обеспечивать жизнедеятельность организма. Длительность митотического периода различна: клетки крови и кожи входят в процесс деления каждые 24 часа, а нейроны способны к репродукции только у новорожденных, а затем вовсе утрачивают способность к размножению.
Существует 2 вида деления — прямое и непрямое . Соматические клетки размножаются непрямым путем, гаметам или половым клеткам присущ мейоз (прямое деление).
Митоз — непрямое деление
Митотический цикл
Митотический цикл включает 2 последовательных этапа: интерфазу и митотическое деление.
Интерфаза (стадия покоя) – подготовка клетки к дальнейшему разделению, где совершается дублирование исходного материала, с последующим его равномерным распределением между новообразованными клетками. Она включает 3 периода:
- Пресинтетический (G-1) G – от английского gar, то есть промежуток, идет подготовка к последующему синтезу ДНК, выработка ферментов. Экспериментально проводилось ингибирование первого периода, вследствие чего клетка не вступала в следующую фазу.
- Синтетический (S) — основа клеточного цикла. Происходит репликация хромосом и центриолей клеточного центра. Только после этого клетка может перейти к митозу.
- Постсинтетический (G-2) или премитотический период — происходит накопление иРНК, которая нужна для наступления собственно митотического этапа. В G-2 периоде синтезируются белки (тубулины) – основная составляющая митотического веретена.
После окончания премитотического периода начинается митотическое деление . Процесс включает 4 фазы:
- Профаза
– в этот период разрушается ядрышко, растворяется мембрана ядра (нуклеолема), центриоли располагаются на противоположных полюсах, формируя аппарат для деления. Имеет две подфазы:
- ранняя — видны нитеобразные тела (хромосомы), они еще не четко отделены друг от друга;
- поздняя — прослеживаются отдельные части хромосом.
- Метафаза – начинается с момента разрушения нуклеолемы, когда хромосомы хаотично лежат в цитоплазме и только начинают двигаться к экваториальной плоскости. Между собой все пары хроматид связаны в месте центромеры.
- Анафаза – в один момент разобщаются все хромосомы и движутся к противоположным точкам клетки. Это короткая и очень важная фаза, поскольку именно в ней происходит точный раздел генетического материала.
- Телофаза – хромосомы останавливаются, снова образуется ядерная мембрана, ядрышка. Посередине образуется перетяжка, она делит тело материнской клетки на две дочерние, завершая митотический процесс. В новообразованных клетках снова начинается G-2 период.
Мейоз — прямое деление
 Мейоз — прямое деление
Мейоз — прямое деление
Существует особый процесс репродукции, встречающийся только в половых клетках (гаметах) – это мейоз (прямое деление) . Отличительной чертой для него является отсутствие интерфазы. Мейоз из одной исходной клетки дает четыре, с гаплоидным набором хромосом. Весь процесс прямого деления включает два последовательных этапа, которые состоят из профазы, метафазы, анафазы и телофазы.
Перед началом профазы у половых клетках происходит удвоение исходного материала, таким образом, она становится тетраплоидной.
Профаза 1:
- Лептотена — хромосомы просматриваются в виде тоненьких ниток, происходит их укорочение.
- Зиготена — стадия конъюгации гомологичных хромосом, как следствие образуются биваленты. Конъюгация важный момент мейоза, хромосомы максимально сближаются друг с другом, чтобы осуществить кроссинговер.
- Пахитена — происходит утолщение хромосом, их все большее укорочение, идет кроссинговер (обмен генетической информацией между гомологичными хромосомами, это основа эволюции и наследственной изменчивости).
- Диплотена – стадия удвоенных нитей, хромосомы каждого бивалента расходятся, сохраняя связь только в области перекреста (хиазмы).
- Диакинез — ДНК начинает конденсироваться, хромосомы становятся совсем короткими и расходятся.
Профаза заканчивается разрушением нуклеолемы и формированием веретена деления.
Метафаза 1 : биваленты расположены посередине клетки.
Анафаза 1 :к противоположным полюсам движутся удвоенные хромосомы.
Телофаза 1 :завершается процесс деления, клетки получают по 23 бивалента.
Без последующего удвоения материала клетка вступает во второй этап деления.
Профаза 2 : снова повторяются все процессы, которые были в профазе 1,а именно конденсация хромосом, что хаотично располагаются между органеллами.
Метафаза 2 : две хроматиды, соединенные в месте перекреста (униваленты), располагаются в экваториальной плоскости, создавая пластинку, названную метафазной.
Анафаза 2: — унивалент разделяется на отдельные хроматиды или монады, и они направляются к разным полюсам клетки.
Телофаза 2
: процесс деления завершается, формируется ядерная оболочка, и каждая клетка получает по 23 хроматиды.
Мейоз – важный механизм в жизни всех организмов. В результате такого деления мы получаем 4 гаплоидные клетки, которые имеют половину нужного набора хроматид. Во время оплодотворения две гаметы образуют полноценную диплоидную клетку, сохраняя присущей ей кариотип.
Сложно представить наше существования без мейотического деления, иначе все организмы с каждым последующим поколение получали бы удвоенные наборы хромосом.
Интерфаза и различные способы деления клеток. Различают два способа деления: 1) наиболее распространенное, полноценное деление - митоз (непрямое деление) и 2) амитоз (прямое деление). Во время митотического деления происходит перестройка цитоплазмы, разрушение оболочки ядра, выявление хромосом. В жизни клетки выделяют период самого митоза и промежуток между делениями, который называют интерфазой. Однако период интерфазы (неделящейся клетки) по своей сущности может быть различным. В одних случаях во время интерфазы клетка функционирует и одновременно готовится к следующему делению. В других случаях клетки переходят в интерфазу, функционируют, но уже не готовятся к делению. В составе сложного многоклеточного организма имеются многочисленные группы клеток, утратившие способность делиться. К числу их относятся, например, нервные клетки. Подготовка клетки к митозу происходит в интерфазе. Для того чтобы представить себе основные черты этого процесса, вспомните строение клеточного ядра.
Основной структурной единицей ядра являются хромосомы, состоящие из ДНК и белка. В ядрах живых неделящихся клеток, как правило, отдельные хромосомы неразличимы, но большая часть хроматина, которую на окрашенных препаратах обнаруживают в форме тонких нитей или зерен различной величины, и соответствует хромосомам. У некоторых клеток отдельные хромосомы отчетливо видны и в интерфазном ядре, например в быстро делящихся клетках развивающегося оплодотворенного яйца и ядрах некоторых простейших. В различные периоды жизни клетки хромосомы претерпевают циклические изменения, которые прослеживаются от одного деления до другого.
Хромосомы во время митоза представляют собой удлиненные плотные тельца, по длине которых можно различать две нити - хроматиды, содержащие ДНК, представляющие собой результат удвоения хромосом. На каждой хромосоме выделяется первичная перетяжка, или центромера. Эта суженная часть хромосомы может быть расположена или посередине, или ближе к одному из концов, но для каждой определенной хромосомы ее место строго постоянно. Во время митоза хромосомы и хроматиды представляют собой туго свернутые спиральные нити (спирализованное, или конденсированное, состояние). В интерфазном ядре хромосомы сильно вытянуты, т. е. деспирализованы, благодаря чему становятся трудноразличимыми. Следовательно, цикл изменения хромосом состоит в спирализации, когда они укорачиваются, утолщаются и становятся хорошо различимыми, и деспирализации, когда они сильно вытягиваются, переплетаются, и тогда уже различить каждую в отдельности становится невозможно. Спирализация и деспирализация связаны с деятельностью ДНК, так как она функционирует только в деспирализованном состоянии. Выдача же информации, образование РНК на ДНК в спирализованном состоянии, т. е. во время митоза, прекращается.
Тот факт, что хромосомы присутствуют в ядре неделящейся клетки, доказывается также постоянством количества ДНК, числа хромосом и сохранением от деления до деления их индивидуальности.
Подготовка клетки к митозу . В течение интерфазы происходит ряд процессов, которые обеспечивают митоз. Назовем главнейшие из них: 1) удваиваются центриоли, 2) удваиваются хромосомы, т.е. количество ДНК и хромосомальных белков, 3) синтезируются белки, из которых строится ахроматиновое веретено, 4) накапливается энергия в виде АТФ, которая расходуется во время деления, 5) заканчивается рост клетки.
Первостепенное значение в подготовке клетки к митозу имеет синтез ДНК и удвоение хромосом.
Удвоение хромосом связано, прежде всего, с синтезом ДНК и одновременно происходящим синтезом белков хромосом. Процесс удвоения продолжается 6-10 часов и занимает среднюю часть интерфазы. Удвоение хромосом протекает так, что каждая старая одиночная цепь ДНК строит себе вторую. Этот процесс строго упорядочен и, начинаясь в нескольких точках, распространяется вдоль всей хромосомы.
Митоз. Фазы митоза

Митоз представляет собой универсальный способ деления клеток растений и животных, основная сущность которого состоит в точном распределении удвоенных хромосом между обеими образующимися дочерними клетками. Подготовка клетки к делению занимает, как мы видим, значительную часть интерфазы, и митоз начинается только тогда, когда подготовка в ядре и цитоплазме полностью заканчивается. Весь процесс подразделяют на четыре фазы. Во время первой из них - профазы - центриоли делятся и начинают расходиться в противоположные стороны. Вокруг них из цитоплазмы образуются ахроматиновые нити, которые вместе с центриолями образуют ахроматиновое веретено. Когда закончится расхождение центриолей, вся клетка оказывается полярной, обе центриоли располагаются у противоположных полюсов, а средняя плоскость может быть названа экватором. Нити ахроматинового веретена сходятся у центриолей и широко располагаются на экваторе, по форме напоминают веретено. Одновременно с образованием в цитоплазме веретена ядро начинает разбухать, и в нем четко выделяется клубок утолщенных нитей - хромосом. На протяжении профазы происходит спирализация хромосом, которые при этом укорачиваются и утолщаются. Профаза заканчивается растворением ядерной оболочки, а хромосомы оказываются лежащими в цитоплазме. В это время видно, что все хромосомы уже двойные.
Затем наступает вторая фаза - метафаза. Хромосомы, расположенные сначала беспорядочно, начинают передвигаться к экватору. Все они обычно располагаются в одной плоскости на равном расстоянии от центриолей. В это время к хромосомам прикрепляется часть нитей веретена, другая же часть их по-прежнему тянется непрерывно от одной центриоли до другой - это опорные нити. Тянущие, или хромосомальные, нити прикрепляются к центромерам (первичным перетяжкам хромосом), но при этом нужно помнить, что как хромосомы, так и центромеры уже двойные. Тянущие нити от полюсов прикрепляются к тем хромосомам, которые к ним ближе. Наступает короткая пауза. Это центральная часть митоза, после которой начинается третья фаза - анафаза.
Во время анафазы тянущие нити веретена начинают сокращаться, растягивая хромосомы к разным полюсам. При этом хромосомы ведут себя пассивно, они, изгибаясь наподобие шпильки, двигаются вперед центромерами, за которые их тянет нить веретена. В начале анафазы снижается вязкость цитоплазмы, что способствует быстрому движению хромосом.
Следовательно, нити веретена обеспечивают точное расхождение хромосом (удвоившихся еще в интерфазе) к разным полюсам клетки.
Завершается митоз последней стадией - телофазой. Хромосомы, приближаясь к полюсам, тесно переплетаются друг с другом. Одновременно начинается их вытягивание (деспирализация), и различить отдельные хромосомы становится невозможным. Постепенно из цитоплазмы образуется ядерная оболочка, ядро разбухает, появляется ядрышко, и восстанавливается прежнее строение интерфазного ядра.

В конце анафазы или в начале телофазы начинается деление цитоплазмы. У клеток животных снаружи в виде кольца появляется перетяжка, которая, углубляясь, разделяет клетку на две меньших размеров. У растений цитоплазматическая оболочка возникает в середине клетки и распространяется к периферии, разделяя клетку пополам. Уже после образования плазматической оболочки у растительных клеток возникает целлюлозная оболочка. Следовательно, в делении клетки активное участие принимает и ядро, и цитоплазма. Ядро содержит уникальные структуры клетки - хромосомы, а ахроматиновое веретено, формирующееся из цитоплазмы, осуществляет их правильное и равное распределение между обеими дочерними клетками.
Продолжительность митоза и интерфазы
Митоз - относительно короткий период в жизни клетки, гораздо дольше длится интерфаза, что видно из таблицы.
В быстро размножающихся клетках митоз может длиться всего несколько минут. Следовательно, продолжительность митоза варьирует от нескольких минут до 2-3 ч. Интерфаза же длится от 8-10 ч. до нескольких суток.
Скорость, с которой протекают отдельные фазы митоза, также различна.